イネ
| イネ | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 成熟期のイネ(長粒種) | ||||||||||||||||||||||||
分類 | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
学名 | ||||||||||||||||||||||||
Oryza sativa | ||||||||||||||||||||||||
和名 | ||||||||||||||||||||||||
| イネ(アジアイネ) | ||||||||||||||||||||||||
| 英名 | ||||||||||||||||||||||||
Rice |
イネ(稲、稻、禾)は、イネ科イネ属の植物[1]。属名Oryza は古代ギリシア語由来のラテン語で「米」または「イネ」を意味する。種小名 sativa は「栽培されている」といった意味である。収穫物は米と呼ばれ、トウモロコシやコムギとともに世界三大穀物の1つとなっている。稲禾(とうか)、禾稲(かとう)などとも呼ばれる。
目次
1 概要
2 原産と伝播
2.1 原産地
2.2 日本への伝播と普及
3 形態
4 分類
4.1 水稲と陸稲
4.2 糯粳性による分類
4.3 早晩性による分類
4.4 有色米
4.5 その他
5 一般的な品種
5.1 日本国内の品種
5.1.1 うるち(粳)
5.1.2 もち(糯)
5.1.3 酒米
5.1.4 観賞用
5.2 日本以外の品種
6 栽培
7 主要病害虫
8 品種改良
8.1 交配法による
8.2 突然変異による
9 モデル植物研究
10 稲に関わる語彙
11 参考画像
12 脚注
13 参考文献
14 関連項目
15 外部リンク
概要
イネ科イネ属の植物には23種77系統が知られている[2]。このうち20種が野生イネであり、2種が栽培イネである[1]。栽培イネの2種とはアジア栽培イネ(アジアイネ、Oryza sativa)とアフリカ栽培イネ(アフリカイネ、グラベリマイネ、Oryza glaberrima)である[1][3]。結実後も親株が枯れず株が生き続ける多年生型と枯れ毎年種子で繁殖する一年生型があるが、2型の変位は連続的で中間型集団も多く存在する[4]。原始的栽培型は、一年生型と多年生型の中間的性質を有した野性イネから生じたとする研究がある[4]。なお、いくつかの野生イネは絶滅したとされている[4]。
アジアイネはアジアのほか、広くヨーロッパ、南北アメリカ大陸、オーストラリア、アフリカで栽培されている[1]。これに対してアフリカイネは西アフリカで局地的に栽培されているにすぎない[1][5]。イネは狭義にはアジアイネを指す[3]。
アジアイネには耐冷性の高いジャポニカ種(日本型)と耐冷性の低いインディカ種(インド型)の2つの系統がある[1][3]。また、これらの交雑による中間的品種群が多数存在する[3]。
- ジャポニカ種(日本型、島嶼型、Oryza sativa subsp. japonica)
日本、朝鮮半島、中国など温帯~亜熱帯の地域で栽培されている[1]。ジャポニカ種は温帯日本型と熱帯日本型(ジャバニカ種)に分けられる[1]。
- 温帯日本型(温帯島嶼型)
- 主に日本や遼寧省で栽培されている[1]。
- 熱帯日本型(ジャバニカ種、熱帯島嶼型、ジャワ型、Oryza sativa subsp. javanica)
- 中国南部などで栽培されている[1]。なお、ジャポニカ種(日本型)、インディカ種(インド型)、ジャバニカ種(ジャワ型)に並列的に分けられることもある[6][5]。
- インディカ種(インド型、Oryza sativa subsp. indica)
インド、スリランカ、台湾南部、中国南部、東南アジアなど熱帯・亜熱帯の地域で栽培されている[1]。インディカ種(インド型)はジャポニカ種(日本型)以上に分化している[1]。
日本の農学者加藤茂苞による研究が嚆矢となったことから、彼の用いた「日本型」「インド型」という呼称が広く使われているが、両者が存在する中国では、加藤の研究以前からこれに相当する「コウ」(粳稻)「セン」(籼稻)という分類が存在している。中国では、淮河と長江との中間地域で両者が混交し、長江以南でセン、淮河以北でコウが優占する。
加藤による命名が象徴するように、それぞれの生態型の栽培地域には耐寒性による地理的勾配が知られている。日本や中国東北部、朝鮮半島では主にジャポニカ種が栽培され、中国南部や東南アジア山岳部ではジャバニカ種が多く、中国南部からインドにかけての広い地域でインディカ種という具合である。ただし、こうした栽培地域の地理的分離は絶対的なものではなく、両方が栽培されている地域も広範囲にわたる。特に雲南からアッサムにかけての地域は、山岳地域ならではの栽培環境の多様性もあり、多くの遺伝変異を蓄積しているとされる。
栽培イネの祖先種とされるのはオリザ・ルフィポゴン(Oryza rufipogon)である[3]。このオリザ・ルフィポゴンの生態型には多年生型と一年生型があり、特に一年生型がOryza nivaraとして別種扱いされることもある。しかし、分子マーカーによる集団構造の解析によっても一年生型と多年生型が種として分化しているという証拠は得られていない[3]。なお、交雑が進んだ結果、今日では栽培イネから遺伝子浸透を受けていない個体群はインドやインドネシアの山岳地帯に残るにすぎない[3]。
イネには亜種や近隣種が多いために予期せぬ雑種交配が起こることがある。特に、亜種の多様な東南アジアにおいては顕著である。日本では雑種交配を防止するため、耕作地周辺を頻繁な雑草刈りで予防している。
栽培イネではなく雑草として生じるものを雑草イネという[7]。こうした雑草イネは生態的および形態的特徴が栽培イネのそれと類似するため、駆除が極めて難しい。雑草イネは水田の強雑草で栽培イネの生育障害、脱粒、収穫種子に赤米として混入し品質低下を引き起こしている[8]。日本では乾田直播栽培で発生しやすい[8]。栽培稲の生産性を落とすだけでなく、栽培イネと交雑することで品質劣化を起こす。東南アジアでは特に顕著で、食用稲の生産性向上の課題となっている。一方で、祖先型野生稲は遺伝資源としての有用性も指摘されており[8]、種子銀行などの施設での保存のほかに、自生地(in situ)での保全の試みもある。
栽培イネ以外ではO. officinalis(薬稲)が救荒植物として利用されることがある。
原産と伝播
原産地
原産地は中国南部の長江下流域とする説、東南アジアとする説がある[9][6][5][10]。考古学の調査によれば、人の手に由る栽培については中国長江流域で約1万2千年前に栽培されていることから、栽培の起源は確定している[3]。ただし、品種その物に関して当時のイネはジャポニカ種とインディカ種に分化する以前の両方の特徴を持っているとの指摘もあり断定はされていない[11]。
日本への伝播と普及
日本国内に稲の祖先型野生種が存在した形跡はなく、揚子江中流地域において栽培作物として確立してから、栽培技術や食文化などと共に伝播したものと考えられている。日本列島への伝播については、いくつかの説があり、概ね以下のいずれかの経路によると考えられている[12]。
- 江南地方(長江下流域)から九州北部への直接ルート、
- 江南地方(長江下流域)から朝鮮半島南西部を経由したルート、
- 南方の照葉樹林文化圏から黒潮に乗ってやって来た「海上の道」ルートである[10]。たふぁし、多様な伝播経路を考慮すべきとの指摘もある[13]。
縄文時代前期の遺跡からは複数のイネ科植物の遺骸であるプラント・オパールが出土している[3]。本格的に稲作が始まった時期は地域によって差があり、最も早いのは九州西北部で弥生時代早期にあたる紀元前9世紀からとされる[7]。ただし、初期の稲作は用水路などの栽培環境が整備された水田ではなく、自然地形を利用する形態で低湿地と隣接する微高地を利用していた[12][14]。
水稲作の日本への伝来は縄文時代後期にあたる紀元前11世紀ごろであるという説もあるが[5]、本格的な栽培が始まるのは近畿地方では紀元前2世紀ごろ、関東地方では2世紀ごろ、本州北端では12世紀ごろ[5]、北海道では明治時代以降である[5]。現在では亜寒帯(亜寒帯湿潤気候)に属する北海道遠別町から熱帯(熱帯雨林気候)に属する沖縄県西表島まで、広い地域で栽培されている。
現在日本で栽培されるイネは、ほぼ全てが温帯日本種に属する品種であるが、過去(2005年2月、岡山市の彦崎貝塚で、縄文時代前期(約6000年前)の土層からイネのプラント・オパールが多量に出土した。同市の朝寝鼻遺跡でも同時期の発見例があり、縄文時代前期から畑作によるイネの栽培が始まっていた可能性が高まった[15])には熱帯日本種(ジャバニカ種)も伝播し栽培されていた形跡があるとされているが、他地域で栽培されたものが持ち込まれた可能性も否定できないとの見解もある[16]。しかし、2008年国立歴史民俗博物館の研究者らは、岡山県彦崎貝塚のサンプルは異なった時代の付着物に汚染され、正確な年代判定が行えていなかったと指摘している[17]。
形態
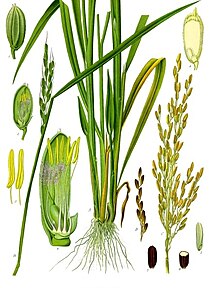
イネの植物図
多くの節をもつ管状の稈を多数分岐させ、節ごとに1枚の細長い肉薄の葉をもつ。また、葉の付け根には葉舌という器官がある。葉には筋状の葉脈にガラス体(プラントオパール)を蓄積させ、そのガラス体の柱(植物骨格組織)を支えにして直立する。薄手の葉が直立する草型のため、密集状態での受光効率が高い。稈は節の詰まったロゼット状になっており、生殖成長期になると徒長して穂を1つつける。栄養成長期と生殖成長期が明確に分かれており、穂を付けるのは稈を増やす時期が終了してからであり、籾が成熟し生殖成長が終わると、ヒコバエが生え再び栄養成長を再開する。
他殖性の風媒花であり、開花前に稈が徒長して穂を草叢から突き出すのは、開花時に花粉を飛ばしやすくするためである。ただし開花前に花粉が熟し、開花時に葯が破裂するため、栽培稲では98%程度が自家受粉する。開花時間は午前中から昼ごろまでの2-3時間と短い。花は、頴花(えいか)と呼ばれ、開花前後の外観は緑色をした籾(もみ)そのものである。籾の先端には、しなやかな芒(ぼう)が発達する。芒は元々は種子を拡散するための器官であるが、栽培上不要なため近代品種では退化している。
農業上、種子として使われる籾は、生物学上の果実である玄米を穎(=籾殻:もみがら)が包んでいるもの。白米は、玄米から糠(ぬか)層、胚など取り除いた、胚乳の一部である。

イネの花

収穫期の穂の拡大写真

籾

イネの刈り取り後に伸びるひこばえ(蘖)




分類
水稲と陸稲
|


元来、イネは湿性植物である[18]。水田で栽培するイネを水稲(すいとう、lowland rice)、耐旱性が強く畑地で栽培するイネを陸稲(りくとう、おかぼ、upland rice)という[1][6][19]。日本では明確に水稲と陸稲が区別されるが[1]、他の国では水稲と陸稲とは明確には区別されていない[1]。
逆転の日本史編集部『日本人のルーツがわかる本によれば、陸稲(熱帯ジャポニカ)に関しては、すでに三内丸山遺跡から「2800年前の『米粒』が見つかっています」と述べ、水稲(温帯ジャポニカ)に関しては「弥生時代頃に、水田耕作技術を持った人々が朝鮮半島の方から日本列島に持って来たもの」と説明している[20]。
水稲には、灌漑稲、天水稲、深水稲、浮稲といった種類がある[18]。水位が著しく上昇して葉が水没するような状況では、節間を急速に伸ばすことで水面から葉を出し、窒息を免れることができる。節間の伸張能力は品種により著しい差があり、数センチから十数メートルまで伸張する品種がある。特に著しく伸張させることができる品種は浮稲(うきいね)と呼称される。
陸稲は水稲に比べて食味の点で劣るとされ、日本においては近年では糯種などが栽培されているにすぎない[19]。
糯粳性による分類
稲の食用部分の主成分であるでんぷんは、分子構造の違いからアミロースとアミロペクチンに別けられる。お米の食感は、両者の含有配分によって大きく異なる。すなわちアミロース含量が少ないお米は加熱時にやわらかくモチモチした食感になり、アミロース含量が多いとパサパサした食感になる。日本人の食文化では、低アミロースのお米を「美味しい」と感じる。この好みは、世界的には少数派となっている。
通常の米は20%程度のアミロースを含んでいるが、遺伝的欠損によりアミロース含量が0%の品種もある。これがモチ性品種である。アミロース含有を0%にする遺伝的欠損をモチ性といい、この特質をもつ作物は稲だけではなく、他にアワ、キビ、ハトムギ、モロコシ、トウモロコシ、オオムギ、アマランサス(けいとう)に見つかっている。これらの作物は世界中で栽培されているにもかかわらず、モチ性品種が栽培されている地域は東南アジア山岳部の照葉樹林帯に限定されている。その特異性から、その地域を「モチ食文化圏」と呼称されることがある。
日本列島自体が西半分を「モチ食文化圏」と同じ照葉樹林に覆われており、またハレの日にもち米を食べる習慣がある(オコワ、赤飯、お餅)ことから、日本文化のルーツ(の一つ)として注目された。
早晩性による分類
早晩性により早稲(わせ)、中稲(なかて)、晩稲(おくて)などに分類される。一般には早生(わせ)、中生(なかて)、晩生(おくて)と表記されるが、イネの場合には特に早稲(わせ)、中稲(なかて)、晩稲(おくて)と表記される。
有色米
黒米、赤米、緑米などを総称して有色米という[10]。野生種に近い米である[10]。

黒米の稲穂
黒米(くろまい、くろごめ)- 果皮の色が黒い米。胚乳部分は普通の米と同様に白い。よってぬか部分を完全にとってしまうと品種の特徴は消えてしまう。
中国や東南アジアでは、一般の食品や酒造原料としても利用されている。一方、日本では希少なため、赤米と共に神事の際に神饌として用いる機会が多い。黒く見える色素はポリフェノールの一種であるアントシアニンに起因しており、非常に濃い紫色である。白米と混ぜて炊飯した時、お米が赤飯のように紫っぽくなるため、紫黒米とも呼ぶ。日本での代表的な品種は、おくのむらさき、朝紫、むらさきの舞、紫黒苑。近年古代米と称し栽培が復活しつつある。また、イタリアではインディカ種白米とジャポニカ種黒米とによる交配研究の結果、1997年にRiso Venereという新たな黒米品種が誕生、現在はヴェルチェッリ県とノヴァーラ県を中心に農作されている。
赤米(あかまい)- 種皮の色が赤い米。胚乳部分は普通の米と同様に白い。種皮(ぬか)の組織が強固で精米によって完全に取り除くことが難しく、赤い玄米状態で利用することになるため赤米と呼称される。祖先型野生稲はすべて赤米であることが知られており、普通稲は種皮の組織をつくる遺伝子の欠損によって生じたものと考えられる。そのため赤米は、ジャポニカ種・インディカ種、陸稲・水稲、粳米・糯米にかかわらず存在する。
- 玄米の表面の層が赤いのはタンニン系の色素を含有しているため。日本では8世紀の頃平城京の木簡から栽培が確認される。また14世紀ころに「大唐米」という長粒種が渡来した。日本での代表的品種は、国司、神丹穂、ベニロマン、紅衣など。江戸時代に関東から西特に薩摩など南九州で多く栽培されていたが、明治以降品種改良米の普及活動により昭和中期には神事用以外は駆逐された。近年古代米と称し栽培が復活しつつある。また、日本には粳米しかなかったが、品種改良により糯米ができた。白米と混ぜて炊くとピンクがかった色になる。
緑米(みどりまい)- 種皮の色が緑色をしたもち米。
その他
- 香り米
- 低アミロース米
- 低グルテリン米
- その他 困窮地域などでの栄養不足を補うために、ビタミンなどを強化した品種もある。
一般的な品種
日本国内の品種
うるち(粳)
うるち米には次のような品種がある。
| 登録番号・名称 | 地方番号(旧系統名) | 交配品種 | 育成機関 | 登録年 |
|---|---|---|---|---|
水稲農林1号 | 北陸4号 | 森多早生/陸羽132号 | 新潟県農事試験場 | 1931年 |
水稲農林20号 | 北海86号 | 水稲農林1号/胆振早稲 | 北海道農業試験場 | 1941年 |
| 水稲農林100号(コシヒカリ) | 越南17号 | 水稲農林22号/水稲農林1号 | 東海近畿農業試験場 | 1956年 |
| 水稲農林108号(ヤマビコ) | 東海7号 | 中京旭/水稲農林22号 | 福井県農事試験場 | 1958年 |
| 水稲農林150号(ササニシキ) | 東北78号 | 奥羽224号/ササシグレ | 宮城県農業試験場 | 1963年 |
日本晴 | 北海86号 | ヤマビコ/幸風 | 愛知県総合農業試験場 | 1963年 |
あきたこまち | 秋田31号 | コシヒカリ/奥羽292号 | 秋田県農業試験場 | 1984年 |
| 水稲農林290号(キヌヒカリ) | 北陸122号 | 収2800/北陸100号/北陸96号 | 北陸農業試験場 | 1988年 |
きらら397 | 上育397号 | 渡育214号/道北36号 | 北海道上川農業試験場 | 1988年 |
| 水稲農林299号(ヒノヒカリ) | 南海102号 | 愛知40号/コシヒカリ | 宮崎県総合農業試験場 | 1989年 |
| 水稲農林313号(ひとめぼれ) | 東北143号 | コシヒカリ/初星 | 宮城県古川農業試験場 | 1991年 |
どまんなか | 山形35号 | 中部42号/庄内29号 | 山形県農業試験場 | 1992年 |
はえぬき | 山形45号 | 庄内29号/秋田31号 | 山形県農業試験場 | 1992年 |
| 水稲農林340号(ほしのゆめ) | 上育418号 | あきたこまち/道北48号/上育397号 | 北海道上川農業試験場 | 1996年 |
つがるロマン | 青系115号 | ふ系141号/あきたこまち | 青森県農業試験場 | 1997年 |
ななつぼし | 空育163号 | ひとめぼれ/空系90242A/空育150号 | 北海道中央農業試験場 | 2001年 |
日本国内における代表的な栽培品種は以下の通り(2009年の収穫量順)[21][22]。
| 順位 | 品種名 | 作付比率(%) | 収穫高 | 収穫高比率(カッコ内順位) |
|---|---|---|---|---|
| 1 | コシヒカリ | 37.3% | 3,094,000t | 36.5%(1) |
| 2 | ひとめぼれ | 10.6% | 842,700t | 10.0%(2) |
| 3 | ヒノヒカリ | 10.3% | 805,300t | 9.5%(3) |
| 4 | あきたこまち | 7.8% | 656,700t | 7.8%(4) |
| 5 | キヌヒカリ | 3.3% | 257,500t | 3.0%(6) |
| 6 | ななつぼし | 3.0% | 200,900t | 2.4%(7) |
| 7 | はえぬき | 2.8% | 258,400t | 3.1%(5) |
| 8 | きらら397 | 2.4% | 167,400t | 2.0%(8) |
| 9 | つがるロマン | 1.6% | 155,300t | 1.8t(9) |
| 10 | まっしぐら | 1.3% | 118,000t | 1.4%(10) |
| 11 | あさひの夢 | 1.2% | 101,100t | 1.2%(11) |
| 12 | 夢つくし | 1.1% | 72,400t | 0.9%(14) |
| 13 | こしいぶき | 1.1% | 89,900t | 1.1%(12) |
| 14 | ほしのゆめ | 1.0% | 67,100t | 0.8%(15) |
| 15 | あいちのかおり | 0.9% | 82,200t | 1.0%(13) |
| 16 | ハナエチゼン | 0.6% | - | - |
| 17 | ハツシモ | 0.6% | - | - |
| 18 | ササニシキ | 0.6% | - | - |
| 19 | 彩のかがやき | 0.6% | - | - |
| 20 | おぼろづき | 0.5% | - | - |
作付高上位10品種で80.4%、上位20品種で88.6%を占める。
1980年代に良食味品種として代表格であったササニシキ・コシヒカリは互いに近縁の関係にあり[23]、両品種以降の後の良食味米は多くはコシヒカリの遺伝子を引き継いでいる。
日本で栽培される稲は遺伝的に近縁の品種が多い。そのため、天候不良や特定の病虫害によって大きく収量を落とす可能性がある。従って、食料の安定生産という観点からより多くの遺伝資源を利用した品種改良が必要である。
もち(糯)
酒米
観賞用
一般的には知られていないが、イネには食用米品種以外に観賞用品種が存在する[24]。観賞用イネは米を収穫することが目的ではなく、鮮やかに染まった葉や穂を鑑賞して楽しむためのイネである。切り花やドライフラワーなどに適している。
また、近年では青森県田舎館村などで取り組まれている田んぼアートなどに用いられている。
- 奥羽観383号
- 奥羽観378号
- 西海観246号
- ゆきあそび
- べにあそび
- あかねあそび
日本以外の品種
- インド
- バスマティ
- タイ
- カオ・ホーム・マリ
- スペイン
- ボンバ米
- イタリア
アルボリオ arborio
カルナローリ carnaroli
ヴィアローネ・ナノ vialone nano
- アフリカ
ネリカ米 NERICA - アフリカの食糧事情改善を目的に開発されたアジアイネ(O. sativa)とアフリカイネ(O. glaberrima)の交雑種。
栽培

田植え後の早苗(初夏))

刈入れ 地方や気候によって時期に違いがある

ハザ干しの様子
イネ(稲)の栽培を稲作(いなさく)という。
栽培する土地を田または田圃(たんぼ)といい、特に水を張っている田を指して水田(すいでん)ともいう。
水田で育成されたものを水稲(すいとう)、畠で育成されたものを陸稲(りくとう・おかぼ)と呼ぶ。日本では、近年では陸稲は少なくなっている。(陸稲は栽培に水が少なくて済むが面積あたりの収穫量が水稲より少ない上に、連作障害が発生する)
水稲は収穫までの間に大量の水を使うが、そのため地力の低下が小さく、永久連作[25](二期作)が可能である。
イネは夏期にある程度高温になる温暖湿潤の気候が適している。しかし寒冷地向け品種の作出と栽培法の確立により、寒冷地での栽培も可能となった。日本では、現在では総生産高のうち、北海道および東北地方が占める割合が最も大きい。東北地方や新潟県の内陸部は夜間の気温が低いためイネの消耗が少なく良食味の米が収穫できるとされ、近年の食味ランキングでは東北地方および新潟県の産米が上位を占めている[26]。しかし、1931年(昭和6年)に並河成資によって世界初の寒冷地用水稲・早稲である農林1号の育成が成功するまでは、現在米どころとされている新潟、山形、秋田など冷涼地の晩稲は「鳥またぎ」とされ、食味では台湾米の比するところではなかった[要出典]。
なお、温帯原産である温帯日本型は本来熱帯気候には適しておらず、温帯に属する日本でも、夏期に猛暑が続くと登熟障害を起こす。近年は地球温暖化に伴い西日本を中心に猛暑日が増え、登熟障害[27]や食味の低下が問題になっており[28]栽培技術による対応[29]だけで無く、耐高温品種の作出も行われている[30]。
主要病害虫
いもち病(稲熱病)- 白葉枯病
- 縞葉枯病
- 立枯細菌病
- 馬鹿苗病
- 籾枯細菌病
- 紋枯病
- イネシンガレセンチュウ
- イネミズゾウムシ
- セジロウンカ
- ヒメトビウンカ
- トビイロウンカ
- ツマグロヨコバイ
- ニカメイチュウ
- イネツトムシ
- フタオビコヤガ
カメムシ類(斑点米を作る)
品種改良
交配法による
コシヒカリに、いもち病の抵抗性遺伝子を連続戻し交配で導入することによって育成されたコシヒカリBLがある。
現在、「新潟県産コシヒカリ」という銘柄は、9割以上がコシヒカリBLという品種群であり、コシヒカリ(品種)とは異なる。
突然変異による
理化学研究所では、重イオンビーム照射により、一般種の1.5倍の耐塩性を獲得した品種の開発に成功[31]。塩害で耕作ができなくなった土地での栽培により、生産可能地域が広がり食糧問題の解決に貢献することが期待される。
モデル植物研究
イネは、生物学や農学において、植物のモデル生物として用いられている。イネは主要穀物の中ではゲノムサイズが小さく(トウモロコシの1/6、小麦の1/40)、穀物の遺伝情報を知る上でモデルとして好適とされる。
農業生物資源研究所がコシヒカリ・ファミリーである「コシヒカリ」「ひとめぼれ」「あきたこまち」「ヒノヒカリ」を分析した結果、6つのDNAを共通して受け継いでいることが判明している。特に興味深いのは明治時代に東西の横綱と称された「亀の尾」、「朝日」のDNAを引いていることが挙げられる[32]。
ゲノム研究所 (TIGR) やイネゲノム研究プログラム (RGP) 国際チームが「日本晴」のゲノムプロジェクトが進行しており、イネゲノムの塩基配列は、2002年12月に重要部分の解読が完了し、2004年12月には完全解読が達成されている[33][34]。先述のようにイネは単子葉植物のモデル生物であり、植物としては双子葉植物であるシロイヌナズナに続いて2番目、単子葉植物としては初めての全ゲノム完全解読となった[1]。
イネは洪水などで水没すると呼吸が出来ず枯れてしまうのが弱点だが、名古屋大学の芦苅基行教授らが水没に耐えられるような茎を伸ばす遺伝子を解明した。他にもいもち病にかかりにくいが食感の悪い「おかぼ」の遺伝子を研究し、食感が失われない「ともほなみ」を2009年に開発している。
稲に関わる語彙
早苗(さなえ) - 女性の名前にも見られる。
稲妻(いなづま)・稲光(いなびかり) - 稲穂の実る時期に雷が多いことから、古来、雷が稲を実らせると考えられていた。
参考画像

頭を垂れる稲穂 ジャポニカ種コシヒカリ系 2007.8.29

10月初旬の稲穂

五円硬貨の表には稲穂がデザインされている。



脚注
- ^ abcdefghijklmnop農業・生物系特定産業技術研究機構編『最新農業技術事典』農山漁村文化協会 p.105 2006年
^ 森島啓子、イネの進化研究を考える 育種学研究 Vol.1 (1999) No.4 P.233-241, doi:10.1270/jsbbr.1.233
- ^ abcdefghi日本作物学会編『作物学用語事典』農山漁村文化協会 p.218 2010年
- ^ abc森島啓子、イネの祖先を探る 日本釀造協會雜誌 Vol.78 (1983) No.9 P.680-683, doi:10.6013/jbrewsocjapan1915.78.680
- ^ abcdef杉田浩一編『日本食品大事典』医歯薬出版 p.9 2008年
- ^ abc『丸善食品総合辞典』丸善 p.411 1998年
- ^ ab日本作物学会編『作物学用語事典』農山漁村文化協会 p.219 2010年
- ^ abc湯陵華、森島啓子、雑草イネの遺伝的特性とその起源に関する考察 育種学雑誌 Vol.47 (1997) No.2 P.153-160, doi:10.1270/jsbbs1951.47.153
^ 平成20年度の主な研究成果 農業生物試験研究所 (PDF)
- ^ abcd『料理食材大事典』主婦の友社 p.307 1996年
^ 王才林、宇田津徹朗ほか、プラント・オパールの形状からみた中国・草鞋全山遺跡(6000年前~現代)に栽培されたイネの品種群およびその歴史的変遷 育種学雑誌 Vol.48 (1998) No.4 P.387-394, doi:10.1270/jsbbs1951.48.387
- ^ ab宇田津徹朗、東アジアにおける水田稲作技術の成立と発達に関する研究 : その現状と課題(日本と中国のフィールド調査から) 名古屋大学加速器質量分析計業績報告書. v.24, 2013, p.113-122, hdl:2237/20151
^ 徐光輝、林留根:長江流域の農耕集落について 龍谷大学国際社会文化研究所紀要 6号,2004, p.57-70, hdl:10519/2341
^ 宇田津徹朗、湯陵華、王才林、鄭雲飛、佐々木章、柳沢一男、藤原宏志:中国・草畦山遺跡における占代水田趾調査(第3報) 日本文化財科学会 学会誌『考古学と自然科学』ISSN 0288-5964 No.43 p.51-64 (PDF)
^ 6000年前の稲作遺物『知恵蔵2014』[信頼性要検証]
^ 平井泰男:大地からのメッセージ(5) 稲作ことはじめ(グラフおかやま1997年8月号より転載) 岡山県古代吉備文化財センター
^ 遠部慎、宮田佳樹ほか、炭素14年代測定に関するサンプリングの実践と課題 国立歴史民俗博物館研究報告 第137集 [共同研究]高精度年代測定法の活用による歴史資料の総合的研究 (PDF)
- ^ ab日本作物学会編『作物学用語事典』農山漁村文化協会 p.220 2010年
- ^ ab杉田浩一編『日本食品大事典』医歯薬出版 p.11 2008年
^ 逆転の日本史編集部『日本人のルーツがわかる本』p83~p85、佐藤洋一郎論文「日本列島に最初に稲作を持ち込んだのは縄文人だった」
^ 農林水産省 - 平成21年産水稲うるち米の作付状況について
^ 農林水産省 - 平成21年産水稲の品種別収穫量
^ 両品種とも明治時代の良食味品種亀の尾と朝日の血統を、水稲農林1号(亀の尾の孫)と水稲農林22号(朝日の孫)を通して引き継いでいる(品種情報:越南17号(コシヒカリ)・品種情報:東北78号(ササニシキ))。
^ 平成17年3月発行のパンフレット「知っていますか?私たちのごはん」(農林水産省総合食料局総務課)より引用。
^ 黒田治之、「わが国果樹栽培技術の課題と展望」日本調理科学会誌 Vol.32 (1999) No.2 p.151-160, doi:10.11402/cookeryscience1995.32.2_151
^ 食味ランキング特A みちのく村山農業協同組合
^ 森田敏、「水稲高温登熟障害の生理生態学的解析」 九州沖縄農業研究センター報告 (52), 1-78, 2009-08, NAID 120005319212
^ 水稲の高温登熟障害の発生要因と対策 (PDF) 和歌山県
^ 高温障害に強い稲の栽培法 全国農業改良普及支援協会 ・(株)クボタ
^ 高温登熟性の高い水稲品種「彩のきずな」の高温条件下における光合成特性 (PDF) 埼玉農総研研報(13)28-33,2014
^ 重イオンビームで新しい植物をつくる理化学研究所ニュース November 2007
^ 【科学】イネの遺伝子研究、ゲノムで加速 国産米の系譜明らかに
^ Pressrease - イネゲノム塩基配列解読記念式典を開催 - 独立行政法人農業生物資源研究所、社団法人農林水産先端技術産業振興センター
^ Pressrease - イネゲノム塩基配列完全解読を達成 - 独立行政法人農業生物資源研究所、社団法人農林水産先端技術産業振興センター
参考文献
- 『新編食用作物』 養賢堂、星川清親著、1980年
- 【子ども向け】「『米』で総合学習みんなで調べて育てて食べよう」シリーズ(全4巻) 金の星社、2002年
関連項目
- 米
- 藁
- 稲作
- 稲妻
田、神田(御田)- 棚田
陸田、乾田、湿田
- 深田
掘り上げ田、掘り下げ田
- 田の神
- 作況指数
- モミラクトンB
外部リンク
(独)農業・食品産業技術総合研究機構 作物研究所 イネ品種特性データベース
Oryzabase(イネ(稲)データベース) ナショナルバイオリソースプロジェクトの一部。
イネにおける生態型と日本品種との系統発生学的研究育種學雜誌 32(4) pp.333-340
イネゲノムプロジェクト
イネの収量を決定する重要遺伝子を同定 -「第2の緑の革命」につながる世界初の成果- 2005/6/24 理化学研究所
完全解読されたイネゲノムの遺伝子3万個を貼付けた「DNAブック(R)」完成 2005/10/19 理化学研究所
イネの収量ホルモンを活性化する遺伝子発見 理化学研究所


